Kompleman Sistemi
Kısaca: Kompleman sistem , Komplement sistemi veya tamamlayıcı sistem, bir canlıdan patojenlerin temizlenmesine yardım eden biyokimyasal bir kaskaddır. Bağışıklık sisteminin bir canlının yaşamı süresindeki gidişlerle değişmeyen ve uyum sağlamayan doğuştan gelen bağışıklık sistemine ait geniş kısımlarıdır. Bununla beraber edinilmiş bağışıklık sistemiyle birlikte işleme konulup, eyleme geçirilebilir. ...devamı ☟
Bu konuda henüz görüş yok.
Doğuştan gelen bağışıklık sistemi
3 yıl önceenflamasyonu iyileştirici bağışıklık sistemi hücreleri, "sitokinler" olarak isimlendirilirler. Bakterileri tanıyacak olan kompleman kaskadlarının aktivasyonu, hücreleri...
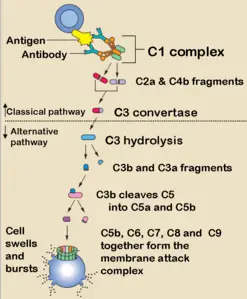
Komplemantasyon
6 yıl önce'kompleman' veya 'komplement', bunun sıfat haline 'komplemanter' veya 'komplementer', bunu yapma özelliğine 'komlemantarite' denir. Kompleman sistemi,...
Bağışıklık Sistemi
3 yıl öncedefensinler olarak adlandırılan antimikrobiyal peptidleri, fagositleri ve kompleman sistemi kapsar. Daha tecrübeli sistemler omurgalıların evrimiyle, nispeten...
Bağışıklık sistemi, Bağışıklık sistemiBiyokimyasal kaskad
6 yıl önceönemli biyokimyasal kaskad tepkimeleri vardır örneğin; koagülasyon kaskadı gibi enzimatik kaskadlar, kompleman sistemi ve sinyal iletimi kaskadları gibi....
Aşırı duyarlılık
3 yıl önceİmmun kompleks, komplemanın C1q komponentini uyararak kompleman sisteminin klasik yoldan aktivasyonunu tetikler. Aktive olan kompleman, eritrositleri farklı...
Aşırı duyarlılık, Akyuvar, Alerji, Alerjik konjonktivit, Anemi, Antijen, Antikor, Astım, BCG, Bazofil, Bağışıklık sistemiMast hücresi
3 yıl öncemediatörleri interstitiuma salarlar. Farklı formlardaki (IgE dışındaki, kompleman sistemi gibi) aktivasyon süreçleri de mümkündür, tanımlanmıştır. Aktivasyon...
Mast hücresi, 1863, 1878, Allerjen, Allerji, Anafilaksi, Antihistamin, Antijen, Antikor, Astım, B hücresiLupus
3 yıl öncezarı glikoproteinleri" gibi insana özgü yıkım artıklarına yapışır ve kompleman yolunu uyarır.Bununla birlikte bölgeye gelen makrofajlar ve lenfositler...
Sistemik lupus eritematozus, Akciğer, Antikor, Artrit, Bağışıklık sistemi, Enflamasyon, Etyoloji, Gastrointestinal sistem, Kalp, Otoantikor, PrevalansTamamlama
3 yıl öncerenk çiftleri Tamamlayıcı sistem kanda bulunan, bağışıklık ile ilgili bir grup protein. Kompleman veya komplement sistemi. Tamamlayıcı tıp, alternatif...