DNA dizilerinin bilinmesi temel biyoloji, biyoteknoloji, adli bilim, tıbbi tanı koyma gibi pek çok sahada vazgeçilmez hale gelmiştir. DNA dizilemesi biyolojik araştırma ve keşifleri çok hızlandırmıştır. Modern DNA dizileme teknolojilerin mümkün kıldığı hızlı DNA dizileme sayesinde İnsan Genom Projesi'nde insan genomunun dizilenebilmiştir. Benzer projelerle pekçok hayvan, bitki ve mikrop genomunun tam dizisi üretilebilmiştir.
İlk DNA dizileri 1970'lerin başlarında üniversite araştırmacıları tarafından iki-boyutlu kromatografiye dayanan zahmetli yöntemlerle elde edilmiştir. Otomatik analizle çalışan boya-tabanlı dizileme yöntemlerinin gelişimiyle DNA dizilemesi çok daha kolaylaşmış ve birkaç büyüklük mertebesi hızlanmıştır.
Tarih
RNA dizilemesi nükleotit dizilemesinin en erken biçimlerinden oldu. RNA dizilemesinin en önemli aşamaları, Gent Üniversitesi'nden (Gent, Belçika) Walter Fiers ve arkadaşlarının, 1972'de Bakteriyofaj MS2'ye ait bir genin, daha sonra 1976'da ise aynı bakteriyofajın tüm genomunun dizisi olmuştur.1970'lerin başlarında hızlı DNA dizileme yöntemlerinin geliştirilene kadar, DNA dizilemesi için çeşitli zahmetli yöntemler kullanılmaktaydı. Örneğin, 1973'te Gilbert ve Maxam, dolaşan benek analiz (wandering-spot analysis) olarak adlandırılan bir yöntemle 24 nükleotitin dizisini yayımladılar.
Sanger ve arkadaşlarının 1975'te geliştirdiği zincir sonlandırma yöntemi kısa sürede en tercih edilen yöntem oldu, göreceli olarak daha kolay ve güvenilir olmasından dolayı.
Maxam - Gilbert dizilemesi
1976-1977'de Harvard Universitesi'nden Allan Maxam and Walter Gilbert, DNA'nın kimyasal modifikasyonu ve ardından onun spesifik bazlarda kesilmesi esasına dayanan bir DNA dizileme yöntemi geliştirdi. Maxam ve Gilbert kimyasal dizileme yöntemi hakkındaki makale, Sanger ve Coulson'un artı-eksi dizilemesi hakkındaki makalesinden iki yıl daha sonra yayınlamasına rağmen, Maxam-Gilbert dizilemesi daha popüler oldu. Bunun nedeni, Maxam-Gilbert yönteminde saflaştırılmış DNA'nın doğrudan dizilenebilmesi, buna karşın ilk Sanger yönteminde tek iplikli DNA üretilebilmesi için okunacak her DNA'nın ayrıca klonlanmasının gerekmesiydi. Ancak, zincir sonladırma yönteminin zaman içinde iyileştirilmesiyle Maxam-Gilbert dizilemesi gözden düştü, zira teknik karmaşıklığı onun standart moleküler biyoloji kit kullanılmasına olanak vermiyordu, ayrıca zararlı kimyasallara gerek gösteriyordu ve ölçeklenmesinde zordu.Bu yöntem DNA'nın 5' ucunun radyoaktif olarak işaretlenmesini (bu tipik olarak gamma-32P ATP kullanılan bir kinaz reaksiyonuydu) ve sonra dizilenecek DNA parçasının saflaştırılmasını gerektirir. Dört farklı kimyasal reaksiyonda (G, A+G, C, C+T) uygulanan kimyasal işlemlerle, moleküllerin ufak bir kısmında, DNA'yı oluşturan dört nükleotit bazdan biri veya ikisinde kesikler meydana gelir. Örneğin, pürinler formik asitle depürine edilir, guaninler (ve bir miktar adeninler de) dimetil sülfat ile metillenir, pirimidinler (C+T) hidrazin ile metillenir. Sadece-C reaksiyonunda, hidrazin reaksiyonuna tuz (sodyum klorür) eklenmesi timinin metillenmesini inhibe eder. Modifiye olmuş DNA'lar sonra sıcak piperidin ile modifiye olmuş bazların poziyonunda kesilir. Modifikasyon yapan kimyasalların konsantrasyonu ayarlanarak DNA molekülü başına ortalama bir modifikasyon olması sağlanır. Böylece bir seri işaretlenmiş DNA parçası elde edilir, bu parçaların bir ucunda radyoaktif işaret, öbür ucunda ise ilk "kesik" yeri olur. Dört reaksiyonda meydana gelen parçalar, denatüran akrilamid jel içinde yanyana elektroforezle ayrıştırılır. Parçaları görselleştirmek için jel bir röntgen filminin üzerine konur, böylece her bir radyoaktif DNA parşasının bulunduğu yere karşılık gelen noktada fotoğraf filmi kararır. Filmde meydan gelen bant serilerinden DNA dizisi çıkarılabilir.
"Kimyasal dizileme" olarak bilinen bu yöntem daha sonradan DNA'ya bağlanan proteinlerin DNA'ya bağlanma yerlerini haritalamak için kullanılan Metilasyon Enterferans Ölçümü'nün temelini oluşturmuştur.
Zincir sonlandırma yöntemleri
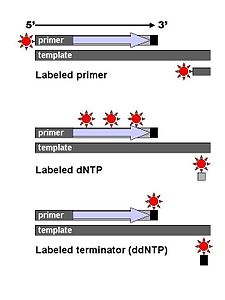
Zincir sonlandırma yöntemi (veya Sanger yöntemi, onu geliştiren Frederick Sanger'e atfen) Maxam ve Gilbert yöntemine kıyasla daha verimli olduğu, daha az toksik kimyasal ve radyoaktivite gerektirdiği için kısa sürede hızla yaygınlaştı. Sanger yönteminin ana ilkesi zincir sonladırıcı olarak dideoksinükleotit trifosfatlar (ddNTP'ler) kullanılmasıdır.Klasik zincir sonladırma yöntemi için tek iplikli bir DNA kalıp, bir DNA primeri, bir DNA polimeraz, normal deoksinükleotitfosfatlar (dNTP'ler) ve DNA zincir büyümesini sonladıran modifiye edilmiş nükleotitler (dideoksiNTP'ler, veya kısaca ddNTP'ler) kullanılır. Bu ddNTP'ler otomatik dizileme makinalarında otomatik olarak tespit edilebilmek için radyokatif veya floresan olarak işaretlenir. DNA numunesi dört ayrı dizileme reaksiyonu için paylaştırılır, bunlarda ortak olarak standart deoksinükleotitler (dATP, dGTP, dCTP ve dTTP) ve DNA polimeraz bulunur. Her reaksiyona dört dideoksinükleotit'ten bir tanesi (ddATP, ddGTP, ddCTP, veya ddTTP) eklenir. İki nükleotit arasında bir fosfodiester bağı oluşması için gerekli olan 3'-OH, dideoksinükleotitlerde bulunmadığı için bunları içeren bir DNA zinciri daha fazla uzayamayaz. Bu nedenle, çeşitli uzunluklarda DNA zincirlerinin oluşumuyla reaksiyon sona erer.
Yeni sentezlenen ve işaretlenen DNA parçaları ısıtılarak denatüre edilir ve jel elektroforeziyle büyüklüklerine göre ayrıştırılır. Dört reaksiyonun (A, C, G ve T) herbirindeki DNA parçaları elektrik alanının etkisiyle jel içinde birbirine paralel ayrı yollardan (bu yollara şerit veya çizgi denir) ilerleyerek ayrıştırılır. Aynı uzunluğa sahip DNA parçları aynı hızda ilerler ve görselleştirildiklerinde (otoradyografi veya mor ötesi ile) birer bant olarak görünürler. Sağdaki resimde jelin üzerine bir röntgen filmi konmuştur, siyah bantlar belli uzunluktaki DNA parçalarına karşılık gelir. Belli bir şeritteki karanlık bant, bir dideoksinükleotitin (ddATP, ddGTP, ddCTP, veya ddTTP) zincire dahi edilmesi sonucu sonlanan bir DNA parçasıdır. Dört şeritteki farklı bantların göreceli konumlarına bakılarak DNA dizisi (alttan yukarı doğru) okunabilir.
ve arkadaşları tarafından geliştirilen bu sistemler otomatik ve yüksek hacimli DNA dizilemesini mümkün kıldı.Zincir sonlandırması DNA dizilemesini son derece kolaylaştırmıştır. Örneğin, ticari olarak elde edilebilen zincir sonladırma kitlerinde dizileme yapmak için gerekli olan reaktantlar, taksim edilmiş ve kullanıma hazır şekilde bulunur. Yöntemin sınırlı kalabileceği durumlar, (1) primerin DNA'ya non-spesifik bağlanıp DNA dizisinin doğru okunmamasına neden olması ve (2) DNA'daki ikincil yapıların dizinin sadakatine etki etmesidir.
Boya sonlandırmalı dizileme
Boya sonlandırmalı dizilemede (İng. Dye-terminator sequencing) zincir sonladırıcı ddNTP'lerin işaretlenir, böylece işaretlenmiş primer yönteminde olduğu gibi dört reaksiyon yerine, dizileme tek bir reaksiyon içinde yürütülebilir. Boya sonladırma dizilemesinde dört dideoksinükleotit bitiricilerin herbiri farklı floresan boyalarla işaretlidir, bunların herbiri farklı dalga boylarında ışık salar.
Daha az işlem gerektirdiğinden ve daha hızlı çalıştığından dolayı, günümüzde otomatik dizileme makinalarının çoğunda boya sonladırma dizilemesi yapılır. Bu yöntemin bazı sınırlamaları vardır, boya işaretli zincir sonladırıcıların DNA parçasına dahil oluşunda boyadan boyaya farklar mevcuttur, bunun sonucu kapiler elektroforezde elde edilen dizi kromatogramındaki pik yükseklikleri ve şekilleri eşitsizlikler gösterir (soldaki şekle bakınız). Modifiye edilmiş DNA polimeraz enzim sistemleri ve DNA'ya dahil olmadaki çeşitliliği azaltan boyalar ile bu problem çözülmüştür. Boya sonladırma yöntemi ile otomatize edilmiş yüksek hacimli DNA dizi analiz makinaları günümüzde dizileme projelerinin büyük bir çoğunluğunda kullanılmaktadır.
Zorluklar
DNA dizilemesinde sık rastlanan sorunlar arasında ilk 15-40 baz dizisinin kötü kaliteli olması, ve 700-900 bazdan sonra dizileme kromatogramının kalitesinin kötüleşmesidir. Baz bildirim (İng. Base calling) yazılımları kalite kırpması (İng.quality trimming) için tipik olarak bir kalite değeri de verir.DNA parçaları dizilemeden önce klonlanmışsa, elde edilen dizide klonlama vektörüne ait kısımlar da olabilir. Buna karşın, PCR-temelli klonlama ve pirodizilemeye dayalı yeni dizileme teknolojilerinde klonlama vektörü olmaz. Ampliseq ve SeqSharp gibi tek adımlı Sanger dizileme (kombine amplifikasyon ve dizileme) yöntemleri sayesinde hedef genler klonlanmadan ve önceden çoğaltılmadan hızlı dizilenebilmektedir.
Mevcut yöntemler tek bir reaksiyonda sadece nispeten kısa (300-1000 nükleotit uzunlukta) DNA parçalarını doğrudan dizileyebilmektedir. Bu limitin üzerindeki DNA parçalarının dizilenmesindeki ana engel, uzunluk farkı bir nükleotit olan büyük DNA parçaları için ayrım gücünün yetersiz olmasıdır.
Otomasyon ve numune hazırlaması
sequencing read display.gif|thumb|right|Bir boya sonlandırma dizileme okumasının başlangıcı.] Otomatik DNA dizileme aletleri (DNA dizileyicileri) bir defada 384 DNA numunesini dizileyebilir ve bu işlemi günde 24 kere tekrarlayabilir. DNA dizileyicileri, kapiler elektroforez kullanarak büyüklük ayrıştırması yapar, boya floresansının tespit ve kayıt eder, ve floresan pik çizim kromatogramı olarak veri çıkarır. Termal döndürücü (İng. thermal cycler) ile dizileme reaksiyonlarının ardından temizleme ve bir tampon çözelti içinde yeniden çözeltme, sonra da dizileyiciye yükleme işlemleri ayrı ayrı yapılır. Düşük kaliteli DNA dizilerinin kırpılmasını otomatik olarak yapabilen çeşitli yazılımlar mevcuttur. Bu programlar her pikteki kaliteyi belirleyip, genelde dizinin sonlarında bulunan düşük kaliteli baz piklerini atarlar. Bu programların doğruluk derecesi insan operatörlerinkinden daha düşüktür ama çok büyük veri kümelerinin otomatik işlenmesi için yeterlidir.Amplifikasyon ve klonal seleksiyon
Büyük ölçekli dizilemenin amacı, kromozom gibi çok uzun DNA parçalarını dizilemektir. Yaygın kullanılan yollardan biri büyük DNA parçalarını restriksiyon enzimleri veya mekanik güçler kullanarak küçük parçalara ayırmaktır. Parçalanmış DNA bir DNA vektörü içine klonlanır ve Escherichia coli içinde çoğaltılır. Her bir bakteriyel koloniden saflaştırılan kısa DNA parçaları ayrı ayrı dizilenir ve tek bir uzun, bitişik dizi şeklinde elektronik olarak birleştirilir.Bu yöntem DNA dizisi hakkında bir ön bilgi gerektirmez ve de novo dizileme olarak adlandırılır. Birleştirilmiş dizideki boşluklar primer yürümesi yapılarak doldurulabilir. Farklı stratejilerin hız ve doğruluk açısından farklı avantajları vardır; saçma yöntemleri (İng. shotgun sequencing) çoğu zaman büyük genomları dizilemekte kullanılır ama elde edilen verinin birleştirmesini yapmak karmaşık ve zordur, özelikle dizi tekrarlarının neden olduğu bölgelerde.
Çoğu dizileme yönteminde bireysel DNA moleküllerinin çoğaltılması için bir in vitro klonlama adımı kullanılır, çünkü moleküler tespit yöntemleri tekil moleküllerin dizilemesine yetecek duyarlıkta değildir. Emülsiyon PCR yönteminde bireysel DNA molekülleri ve primer kaplı boncuklarlar, yağ fazı içinde bulunan sulu damlacıklar içinde izole edilir. Polimeraz zincir tepkimesi (PCR) ile her boncuk DNA molekülünün klonal kopyaları ile kaplanır, sonra bu boncuklar daha sonra dizilemede kullanılmak üzere sabitlenir (immobilize edilir). Emülsiyon PCR Marguilis ve arkadaşları (454 Life Sciences tarafından ticarileştirilmiştir), Shendure ve Porreca ve arkadaşları (Poloni dizilemesi olarak bilinir) ve SOLiD dizilemesi (Agencourt tarafından geliştirilmiştir, artık Applied Biosystems bünyesindedir).
In vitro klonal çoğaltma için kullanıla bir diğer yöntem köprü PCR (İng. bridge PCR) olarak adlandırılır, bunda DNA parçaları katı bir yüzeye bağlanmış primerler üzerinde çoğaltılır (bu yöntem Illumina Genome Analyzer makinasında kullanılır). Tek molekül yöntemleri (Stephen Quake'in laboratuvarında geliştirilen ve daha sonra Helicos tarafından ticarileştirilen) bir istisnadır: bu yöntemde parlak floresan boyalar ve lazer uyarımı kullanılarak bir yüzey üzerine sabitleştirilmiş bireysel DNA moleküllerindeki baz ekleme olayları tespit edilir, böylece moleküler çoğaltma gereği ortadan kalkar.
Yüksek hacimli dizileme
Düşük masraflı dizilemeye olan büyük talep, yüksek hacimli dizileme teknolojilerinin geliştirilmesine yol açmıştır. Bu teknolojilerde dizileme süreci paralelleştirilerek binlerce veya milyonlarca dizi aynı anda üretilir. Yüksek hacimli dizileme teknolojilerinin amacı, standart boya sonlandırma yöntemleri ile mümkün olan DNA dizileme masrafını azaltmaktır.Lynx Therapeutics'in Kitlesel Paralel İmza Dizilemesi
"Gelecek kuşak" dizileme teknolojilerinin ilki olan Kitlesel Paralel İmza Dizilemesi (İng. Massively Parallel Signature Sequencing veya MPSS), 1990'larda Lynx Therapeutics'te geliştirilmiştir. Bu şirket 1992'de Sydney Brenner ve Sam Eletr tarafından kurulmuştur. MPSS, boncuk temelli bir yöntemdi, bir adaptör ligasyonu ve onu takip eden bir adaptör deşifrasyon adımı içeriyordu, ve dizi dört nükleotitlik birimler halinde okunuyordu. Bu yöntem diziye özgü taraflılığa yol açıyordu ve spesifik dizilerin kaybına yol açıyordu. Teknoloji çok karmaşık olduğu için MPSS sadece Lynx Therapeutics tarafından bir hizmet olarak sunulmaktaydı ve dizileme makinası satılmıyordu. Daha sonradan Solexa ile şirket evliliği olunca bu teknoloji, sentez yoluyla dizilemenin gelişmesine yol açtı, çok daha basit olan bu yeni yaklaşım sayesinde MPSS kulanılmaz oldu. Ancak, MPSS çıktısının esas özellikleri daha sonra gelen "gelecek kuşak" (İng. "next gen") veri tiplerine çok benzemekteydi, yüzlerce bin sayıda kısa diziler üretmesi de dahil olmak üzere. MPSS durumunda, bu kısa diziler tipik olarak cDNA dizilemesinde, gen ifade düzeylerini ölçmek amacıyla kullanılıyordu. Lynx Therapeutics 2004'te Solexa ile birleşti ve bu şirket de daha sonra Illumina tarafından satın alındı.
Poloni dizilemesi
Harvard'da George Church'ün laboratuvarında geliştirilen poloni dizilemesi, 2005'te tüm bir genomu dizilemek için kullanılan ilk gelecek-kuşak dizileme sistemleri arasındaydı. In vitro Eşli etiket ("paired-tag") kitaplığı, emülsiyon PCR'si, otomatik bir mikroskop ve ligasyon-temelli dizileme kimyasını birleştirerek bir E. coli genomunu %99.9999'dan yüksek bir doğruluk üretebilmiş ve bunun maliyetini Sanger dizilemesinin yaklaşık 1/10'una getirmiştir. Bu teknoloji önce Agencourt Biosciences şirketine lisanslanmış, daha sonra ana şirketten ayrılan Agencourt Personal Genomics'e aktarılmış ve en nihayet Applied Biosystems'in SOLiD dizileme platformuna dahil edilmiştir.
454 pirodizilemesi
Pirodizilemenin paralelleştirilmiş bir versiyonu 454 Life Sciences tarafından geliştirilmiştir. Bu yöntemde bir yağ solüsyonu içindeki su damlacıklarında yer alan DNA, PCR ile çoğaltılır (emülsiyon PCR'si). Tek bir DNA kalıbının tek bir primer kaplı boncuğa bağlı olduğu her damlacık, sonra klonal bir koloni oluşturur. Dizileme makinası çok sayıda pikolitre hacimli kuyulara sahiptir, bunların her biri tek bir boncuk ve dizileme enzimleri içerir. Pirodizilemede lüsiferaz enzimi kullanılır, bu enzimin ürettiği ışık ile büyümekte olan DNA'ya eklenen herbir nükleotitin tespit edilir. Veriler birleştirilerek dizi okumaları üretilir. Bu teknoloji orta uzunlukta diziler üretir, baz başına maliyeti Sanger dizilemesi ile SOLiD arasında yer alır.
SOLiD dizilemesi
Applied Biosystems'in SOLiD teknolojisinde ligasyon yoluyla dizileme yöntemi kullanılır. Burada, belli uzunluktaki tüm olası oligonükleotitlerin bir karışımı dizilenen konuma göre işaretlenir. Oligonükleotitler tavlanır ve ligasyona uğrar; DNA ligazın birbiriyle eşleşen dizileri bağlama tercihi nedeniyle, o pozisyon için bilgilendirici bir sinyal elde edilir. Dizilemeden önce DNA emülsiyon PCR ile çoğaltılır. Elde edilen boncukların her birinde aynı DNA'nın kopyaları bulunmaktadır, bu boncuklar bir cam lamın üzerine yerleştirilir. Sonuç, Illumina dizilemesine benzer sayı ve uzunluklarda dizilerdir.
DNA nanotop dizilemesi
DNA nanotop dizilemesi Complete Genomics şirketi tarafından geliştirilmiş bir dizileme teknolojisidir. Yöntem, dönen çember ikileşmesi kullanarak genomik DNA'dan elde edilmiş küçük parçaları çoğaltıp bunlardan DNA nanotopları meydana getirmektedir. Ligaz yoluyla dizileme ile nükleotit dizisi belirlenir. Bu yöntem bir defada çok sayıda DNA nanotoplarının dizilenmesini sağlar ve diğer gelecek-kuşak dizileme platformlarına kıyasla daha düşük rektant masrafı vardır. Ancak, herbir DNA nanotopundan sadece kısa diziler elde edilir, bu yüzden bu kısa dizilerin bir referans genomu üzerinde haritalanması zordur. Bu teknoloji çeşitli genom dizileme projelerinde uygulanmıştır.
Gelecek yöntemler
Hibridizasyon yoluyla dizileme DNA mikrodizilimi kullanan non-enzimatik bir yöntemdir. Dizisi belirlenecek olan bir DNA havuzu, floresan işaretlenip üzerinde bilinen diziler bulunan bir mikroçipe hibiridize edilir. Çip üzerinde belli bir noktada kuvvetli hibridizasyon sinyali olması, dizilenmekte olan DNA'da o dizinin bulunduğunu gösterir. Kütle spektrometresi zincir sonlandırma reaksiyonlarında meydana gelen DNA parçaları arasındaki kütle farklarının belirlenmesi için kullanılabilir.Halen geliştirilmekte olan DNA dizileme yöntemleri arasında DNA polimerazın işaretlenmesi, DNA bir nanopordan geçerken dizisinin okunması, ve ağır elementlerle (halojenler gibi) nükleotitleri işaretleyerek uzun DNA parçalarıdaki (>5,000 bp) bireysel nükleotitlerin yerlerini tespit etmeyi amaçlayan mikroskopi temelli teknikler (atomik kuvvet mikroskopu veya transmisyon elektron mikroskopisi ).
Ekim 2006'da X Prize Foundation (X Ödül Vakfı) tüm genom dizileme teknolojilerinin geliştirilmesine önayak olmak için Archon X Ödülü'nü ilan etti. Bu ödül "100 insan genomunu 10 gün veya daha kısa bir zamanda, dizilenen her 100.000 bazdan en fazla 1 hatalık bir doğrulukla, genomdaki dizilerin en az %98'ini içeren diziler üreten ve genom başına $10,000 (US) maliyete bunu yapabilen ekibe" $10 milyon ödül vadetmektedir.
Her sene NHGRI genomiks sahasında yeni araştırma ve geliştirmeler için fonlar vermektedir. 2010 ve 2011'de desteklenen çalışmalar mikroakışkan, poloni ve ağır baz dizileme yöntemleri ile ilgiliydi.
DNA dizilemesinde önemli aşamalar
- 1953: DNA çifte sarmal yapısının keşfi.
- 1972: Rekombinant DNA teknolojisinin keşfi. Bunun sayesinde tanımlı DNA parçalarının izolasyonu mümkün hale gelmiştir. Bundan evvel dizilenebilen numeneler sadece bakteriyofaj veya virüs DNA'larıydı.
- 1977: İlk dizilenen DNA genom bacteriophage φX174.
- 1977: Allan Maxam ve Walter Gilbert "Kimyasal yıkım ile DNA dizilemesi" adlı makalelerini yayımladılar.
- 1984: Britanya'da Tıbbi Araştırma Derneği araştırmacıları Epstein-Barr virüsünün DNA dizisini (170 kb) yayımladılar .
- 1986: California Institute of Technology'de Leroy E. Hood'un laboratuvarı ve Smith ilk yarı otomatik DNA dizileme makinasını ilan ettiler.
- 1987: Applied Biosystems ilk tamamen otomatik otomatik dizileme makinası olan model ABI 370'ı pazarladı.
- 1990: ABD Milli Sağlık kurumu (NIH), Mycoplasma capricolum, Escherichia coli, Caenorhabditis elegans, ve Saccharomyces cerevisiae için ilk büyük ölçekli denemelere başladı (baz başına US$0.75 masraf ile).
- 1991: ifade edilmiş dizi etiketlerinin (İng. expressed sequence tag) dizilenmesi Craig Venter'in laboratuvarında başladı, insan genomunun kodlayıcı kısmını elde etmek amacıyla.
- 1995: The Institute for Genomic Research (TIGR)'da Craig Venter, Hamilton Smith, ve arkadaşları doğada yaşayan bir organizmanın (Haemophilus influenzae bakterisinin) ilk genom dizisini yayımladılar. Dairesel kromozomda 1.830.137 baz bulunmaktaydı ve Science dergisinde yayımlanması tüm-genom saçma dizilime (whole-genome shotgun sequencing) yönteminin ilk kullanımı olmuştur, bu yöntem ile, daha evvelki dizileme projelerinde kullanılan baştan bir haritalama aşaması bertaraf edilmiştir.
- 1996 Stokholm'daki Kraliyet Teknoloji Enstitüsü'nden Nyrén] ve öğrencisi Mostafa Ronaghi pirodizileme yöntemini yayınladılar
- 1998 University of Washington'dan Phil Green ve Brent Ewing, dizileyici veri analizi için
“phred”yayımladılar. - 2000: Lynx Therapeutics, paralelleştirilmiş, adaptör/ligasyon aracılıklı, boncuk tabanlı bir dizileme teknolojisi olan "MPSS"yi yayımladı ve pazarlayıp "gelecek kuşak" dizileme akımını başlattı.
- 2001: insan genomunun taslak sürümü yayımlandı.
- 2004: 454 Life Sciences pirodizilemenin paralelleştirilmiş bir versiyonunu pazarladı. Makinalarının ilk sürümü, otomatik Sanger dizilemesine kıyasla dizileme masraflarını 6-kat azaltmakta, ve MPSS'ninkinin ardından "gelecek kuşak" dizileme makinalarının ikincisiydi.
Kaynaklar
Sekveneerimine